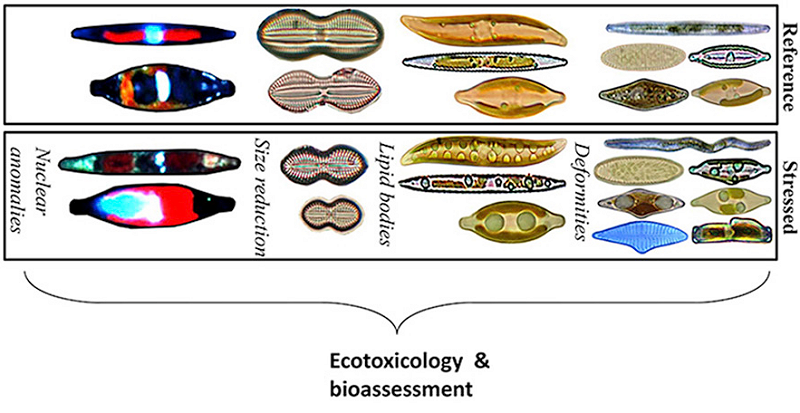
Конспект статьи Л. В. Белоусова (2009) «Морфогенез, морфомеханика и геном» с моими комментариями
* * *
- Морфогенез — это когда организм в процессе развития меняет форму и образует новые структуры.
- Любое развитие организма является морфогенезом.
- Механизмы морфогенеза до сих пор не понятны.
- Преформация — гипотеза о том, что все структуры организма размечены изначально. Она не верна.
- Шаги морфогенеза не записаны в ДНК. Зная полный геном, невозможно предсказать этапы морфогенеза и во что превратится организм в финале.
- Скорее всего геном выступает в роли пассивного инструмента. Его использует организм для производства нужных ему молекулярных структур — в частности, белков. Но тогда что является организмом?
- Морфогенез — это процесс самоорганизации.
* * *
Морфогенез — это развитие организма, механизмы которого не понятны
Есть очень много работ, в которых описан морфогенез того или иного органа или организма в целом, и есть работы в которых высказываются гипотезы о движущих силах морфогенеза, но общепринятой теории морфогенеза нет.
.conspectus Морфогенез в широком понимании — это образование новых форм и структур (от внутриклеточных до макроскопических) в ходе эмбрионального развития организмов.
Морфогенез текстуры панциря диатомовых не соответствует второй части определения, так как у микроводорослей нет эмбрионального развития. Но он соответствует первой части, потому что в процессе морфогенеза организм строит панцирь, то есть создает новые структуры.
Внутриклеточный морфогенез обычно называют цитодифференцировкой. С морфогенезом связан буквально каждый процесс индивидуального развития.
Обычно говорят про морфогенез, а не цитодифференцировку панциря диатомовых.
Морфогенез представляет собой наиболее подробный и в то же время упорядоченный процесс самоусложнения из всех происходящих как в живой, так и в неживой природе.
Общепринятой теории морфогенеза до сих пор не существует.
Мы не можем ответить, почему по ходу развития одни пространственные структуры сменяются строго определенными другими или почему в некоторых случаях закономерные финальные структуры возникают из вариабельных и даже беспорядочных, находящихся на грани хаоса.
Как правило, вопрос [о причинах и механизмах регулирующих морфогенез] так и не ставиться. Подавляющее большинство исследователей вообще не рассматривает морфогенез как самостоятельную проблему. Более того, пока считается, что такой проблемы вообще не существует.
Гипотеза об изначальной разметке структур организма не верна
Преформация — учение, утверждающее, что морфогенез как процесс самоусложнения вообще фиктивен, а все структуры организма пространственно размечены «изначально», в некоторый загадочный начальный момент времени.
Есть аналогичная концепция претекстуры диатомовых (diatoms prepattern) — некого шаблона, на который осаждаются частички кремния, что формирует текстуру.
Отсюда ↓
«How does an amorphous substance produce such a variety of organized shapes? Two major approaches have been followed: either (i) the pattern forms spontaneously out of silica by diffusion limited aggregation (DLA) or (ii) there is a prepattern of something else onto which or within which the silica precipitates (which begs the question as to what causes that pattern)» .
«If silica is ’merely’ precipitated onto an already existed scaffold (prepattern), as some would hypothesize, then we ought to be able to construct artificial scaffolds for silica precipitation».
«The compustat was conceived as a means of pushing diatom morphology to match any preconception we might have. It would work by visually scanning all diatoms in a small growth chamber with a digital camera attached to a motorized microscope, and matching the observed patterns against an ideal ’template’».
Преформация долгое время господствовала в эмбриологии. Хотя Г. Дриш в начале XX века открыл эмбриональные регуляции и строго доказал отсутствие изначальной «разметки», отдельные фрагменты этого воззрения существуют до сих пор.
Эмбриональнае регуляция — восстановление нормальной, геометрически правильной и полной структуры организма, несмотря на удаление, добавление или перемешивание его части. Тогда значение каждого элемента системы есть функция его положения.
Для объяснения морфогенеза используют: позиционную информацию, морфогенетическое поле, мозаичность развития, недришевские регуляции, внеклеточный матрикс, механические напряжения.
Геном не содержит алгоритма морфогенеза
К ним [воззрениям о преформации] относится утверждение, что
сам по себе развивающийся организм не обладает самостоятельной динамикой, а лишь пассивно воспринимает предельно детализированную информацию, записанную на матрице. Таковы представления о том, что морфогенез и вообще все процессы развития «запрограммированы генетически».
Доведение этого утверждения до логического предела означало бы, что
если мы располагаем полной информацией о структуре генома особи, мы сможем предсказать ее морфогенез.
Вся совокупность имеющихся фактов показывает несостоятельность этого утверждения.
См. например, «Пути „гены-признаки“ неисповедимы» и «От генетики к эконике». Здесь речь идет об эпигенетике, когда геном задает архитектуру системы, а некоторая (неизвестная) надгенетическая среда и внешние факторы формируют программный код, что вместе создает организм.
Морфогенез лишь в малой степени видоспецифичен.
Наиболее фундаментальные процессы [морфогенеза] протекают сходным образом у генетически весьма различных организмов
Значит алгоритм морфогенеза записан на надгенетическом уровне. Если он вообще где-то записан, а не создается по мере «выполнения». Ну как самораспаковывающийся код самовыполнемой программы.
Геном и морфогенез — сущности совершенно разного порядка
Гены всегда рассматриваются как статические дискретные факторы, тогда как морфогенез — это разворачивающийся в пространстве-времени непрерывный (континуальный — не расчленяемый на отдельные признаки) процесс. Понятно, что прямая проекция одного на другое невозможна.
Даже если принять, что каждый шаг морфогенеза связан с активацией или репрессией отдельных генов (на самом деле это не так), то пространственно-временное расписание активации и репрессии генов должно определяться не ими самими, а вне- (эпи-) генетическими факторами, прямо или косвенно связанными с морфогенезом.
Интересно, как исследователи соотносят гены и морфогенез диатомовых?
Исследования последних десятилетий говорят о существенно большем:
оказалось, что одни и те же (точнее гомологичные) наборы генов, белковые факторы или сигнальные каскады обеспечивают совершенно разные морфогенетические процессы
как у разных видов, так и на разных стадиях развития одного и того же вида.
Все-таки, геном задает конечную конструкцию организма или нет? Или он задает приблизительный финальный вид, который может варьировать в определенных пределах?
С другой стороны
гомологичные эмбриональные структуры у близких видов могут иметь негомологичное генетическое обеспечение.
Это было ясно уже Н. И. Вавилову и позже было подтверждено.
В статье по ссылке не разбирался.
Зная, какой ген экспрессируется или какой сигнальный каскад в данный момент развития и в данной локализации работает, мы ничего не можем сказать о том, какой морфогенетический процесс состоится.
Очевидно, что природа располагает весьма ограниченным набором генетических факторов и сигнальных путей, которыми организм распоряжается согласно плану своего развития как необходимыми, но не содержащими информацию орудиями. Так молоток необходим, чтобы забить гвоздь, но не он определяет, где и когда гвоздь будет забит.
Наглядная аналогия генома и молотка.
То есть не геном управляет конечным устройством организмов, а организм распоряжается геномом, для построения себя, согласно плану своего развития. То есть план развития записан не в геноме.
То есть трудно избежать парадоксального утверждения, что генетические факторы вовсе не содержат в себе сколько-нибудь однозначно информации о развитии.
Информация о развитии [алгоритм морфогенеза] возникает по ходу дела в самой развивающейся системе
Это вплотную подводит нас к представлению о морфогенезе как о процессе самоорганизации.
—-
Если вам вдруг непреодолимо захотелось меня поблагодарить, переведите мне 250 руб. на ароматный капучино с маленьким пироженным.



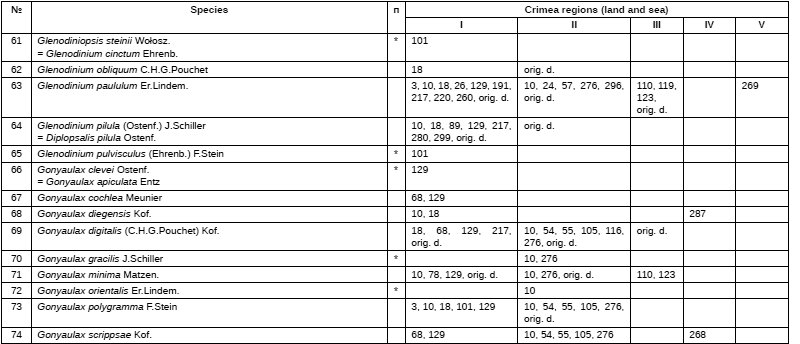
---title.png)
---title.png)




.png)
.png)